
Garden and Plate
The Molecular Biology of Nutrition



Cellular respiration is the flip side of photosynthesis. Photosynthesis captures the energy of the sun in the chemical bonds of ATP and NADPH, and then stores it in the chemical bonds of carbohydrates. Respiration transfers the energy stored in the chemical bonds of carbohydrates back into ATP and NADPH. These molecules then release the energy in just the right amounts to enable the many chemical reactions required in the cell. Cellular respiration is a process that's divided into four main stages: Glycolysis, Pyruvate Decarboxylation, the Citric Acid Cycle, and the Electron Transport Chain.
The first stage of cellular respiration is a ten-step process called glycolysis. It starts with a glucose molecule and ends with two pyruvate, two ATP and two NADH molecules.
Glycolysis evolved earlier than the other stages of cellular respiration, but is less efficient. It doesn't require free oxygen, so it takes place in the cytosol of all living organisms. The later stages were added to cellular respiration for some organisms when free oxygen made better methods (like the electron transport chain) possible.
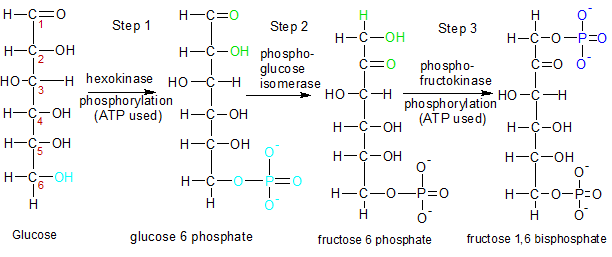
Figure One - Steps 1 through 3: Energy from two ATP molecules is added now so that four ATP and two NADH molecules can be produced in steps 6-10. |
STEP 1: The hexokinase enzyme starts glycolysis when it phosphorylates a molecule of glucose. It aquires the phosphate group from an ATP molecule, converting it to ADP in the process. Glucose is routinely phosphorylated when it enters a cell because the attached phosphate group keeps it from exiting the cell. The changes made in step one are highlighted in light blue and the carbons are numbered with red digits. Now that it's glucose 6-phosphate, it's ready for the second step of glycolysis.
STEP 2: Phosphoglucose isomerase converts glucose 6-phosphate into fructose 6-phosphate by moving the carbonyl oxygen (C=O) from carbon 1 to carbon 2. The atoms involved in the move are green.
STEP 3: Phosphofructokinase attaches a phosphate group from another ATP molecule to carbon 1 on fructose 6-phosphate, creating a molecule of fructose 1,6 bisphosphate. Note that the numbers in all of these molecular names refers to the carbons that have phosphate groups attached.
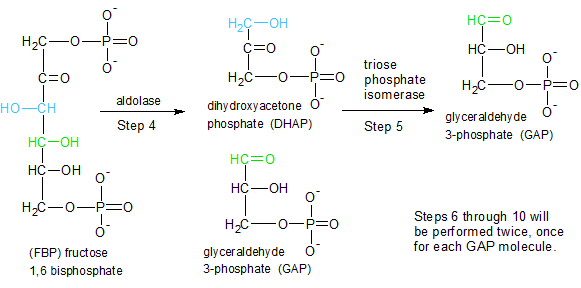
Figure 2 - Steps 4 and 5: The energy laden molecule produced in steps 1-3 is split in preparation for harvesting the energy in steps 6-10. |
STEP 4: The aldolase enzyme splits fructose 1,6 bisphosphate into two molecules. Glyceraldehyde 3-phosphate (GAP) is ready to proceed to step six, but the dihydroxyacetone phosphate (DHAP) molecule needs more work done on it.
STEP 5: The trios phosphate isomerase enzyme converts dihydroxyacetone phosphate (DHAP) into Glyceraldehyde 3-phosphate (GAP) by moving the carbonyl oxygen (C=O) from carbon 2 to carbon 1. We now have two GAP molecules.
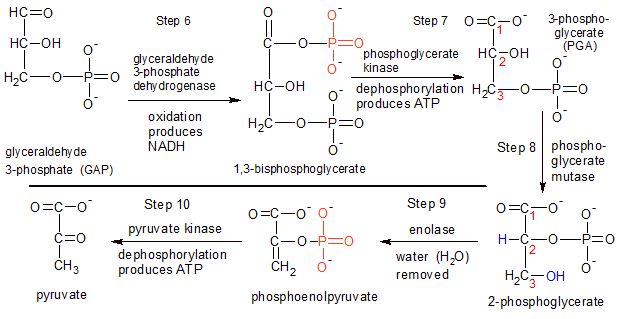
Figure 3 - Steps 6 through 10: This is where the payoff occurs. For an investment of one glucose and two ATP molecules, we get four ATP, two NADH, and two pyruvate molecules. |
NOTE: Steps 6-10 are completed twice, once for each of the two GAP molecules produced at the end of steps 4 and 5.
STEP 6: Glyceraldehyde 3-phosphate dehydrogenase oxidizes the GAP molecule. It binds a free-floating molecule of inorganic phosphate (HPO4-2) to the carbon-1 atom on GAP, forming a C-O bond between them. The hydrogen (H) atom attached to GAP's carbon 1 atom is transferred to a free-floating molecule of NAD+, creating a molecule of NADH+. The H atom attached to the inorganic phosphate molecule donates it's electron to NADH+, creating NADH. The remaining H+ ion is discarded.
STEP 7: Phosphoglycerate kinase transfers a phosphate group (PO3-2) from the carbon-1 atom on 1,3 bisphosphoglycerate to ADP, producing a molecule of ATP. Bisphosphoglycerate is converted into 3-phosphoglycerate in the process.
STEP 8: Phosphoglycerate mutase transfers the remaining phosphate group on 3-phosphoglycerate from carbon-3 to carbon-2. Because the phosphate group is now on carbon-2, the new molecule is called 2-phosphoglycerate. I've placed red numbers next to the carbons on both molecules as a reference.
STEP 9: The enolase enzyme removes a hydroxyl (OH) group from carbon-3, and a hydrogen (H) atom from carbon-2 (both shown in blue). These two fragments combine into a water molecule, which is discarded. Removing these fragments shifts more of the free energy in the new phosphoenolpyruvate molecule to the phosphoral group, setting it up for step ten.
STEP 10: Pyruvate kinase transfers the remaining phosphate group from phosphoenolpyruvate to ADP, creating another molecule of ATP. Phosphoenolpyruvate becomes pyruvate as it loses it's phosphate group.
Final Note: Steps 6-10 produced one NADH and two ATP for each GAP molecule. We started with two GAP molecules at the end of step five, so we really produced two NADH and four ATP. Steps 1-3 used two ATP molecules, so we end up with a total net gain of two NADH and two ATP, with two molecules of pyruvate left over. Glycolysis isn't very productive compared to the later stages of cellular respiration, as we shall see. Pyruvate is still chock full of untapped energy as it moves into the next stage.
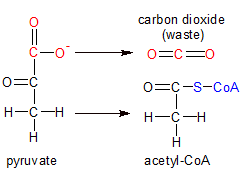
Figure 4 - Pyruvate is converted to acetyl, then combined with co-enzyme A to form acetyl-CoA. |
Glycolysis occurs in the cytosol of every cell, while the later stages of cellular respiration take place in the cytosol of procaryotic cells and the mitochondria of eucaryotic cells. Pyruvate, the end product of glycolysis, is transported into the mitochondria where it's de-carboxylated into an acetyl group and then combined with Coenzyme-A into acetyl-CoA (see figure 4).
A complex of three kinds of enzymes that are collectively called the pyruvate dehydrogenase complex perform these reactions. The three enzymes are pyruvate decarboxylase, lipoamide reductase-transacetylase and dihydrolipoyl dehydrogenase. The complex has multiple copies of each enzyme.
Pyruvate is decarboxylated when the enzyme complex removes carbon-1 from pyruvate, along with the two oxygens attached to it. These are discarded as a molecule of carbon dioxide (CO2). An acetyl group is all that's left of the pyruvate molecule. The enzyme complex then attaches carbon-2 (now carbon-1) of the acetyl group to a sulphur atom on co-enzyme A, forming acetyl-CoA. As part of this process, one molecule of NAD+ is converted to NADH. Acetyl-CoA then transports it's two newly aquired carbons to the Citric Acid Cycle, where they provide the raw material for energy production in the next stages of cellular respiration.
The structures of the molecules involved in the Citric Acid Cycle and the changes they go through are shown in this video produced by Garland Science. It's included as part of the DVD that accompanies their textbook, Molecular Biology of the Cell, 5th ed.
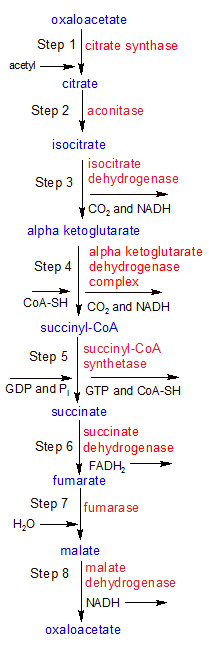
The third stage of respiration is called the citric acid cycle (aka the Krebs or TCA cycle). The citric acid cycle takes place in the mitochondria of eucaryotic cells and starts with one oxaloacetate and one acetyl-CoA molecule. By the end of each cycle it has discarded two molecules of CO2 as waste, created five energy carrier molecules (one each of GTP and FADH2, plus three NADH), and ended up with it's original molecule of oxaloacetate.
The oxaloacetate molecule will be used to start the cycle all over again with a new acetyl-CoA molecule. The GTP transfers it's terminal phosphate group to ADP, creating a molecule of ATP. The other four energy molecules move on to stage four, where they donate their high-energy electrons to the electron transport chain so that more ATP can be created. See the flowchart on the far right for more details.
The Electron Transport Chain from Garland Science.
The fourth and final stage of respiration is called "oxidative phosphorylation". This is the only stage that uses oxygen in the form of O2 gas (making it aerobic), and is the stage where most of the ATP is produced.
The FADH2 and NADH molecules that were created during the citric acid cycle transfer their newly aquired electrons to a series of specialized molecules. These molecules are collectively referred to as the "electron transport chain". Each time an electron is transferred to the next molecule in the chain, it moves to a lower energy level (like going down stairs). The electron moves down the chain because the next molecule over is always more electronegative than the last. This not only attracts the electron, but releases energy as the electron moves into a "lower-energy" position. The electron is finally passed on to a highly electro-negative O2 molecule that uses the electrons and nearby protons (H+ ions) to form water (H2O).
The energy released by the electrons motion down the chain is used to pump protons across the inner mitochondrial membrane to an area between the inner and outer membranes. The protons eventually pile up between the two membranes and diffuse back into the center of the mitochondria through an embedded protein called ATP synthase. This protein uses the energy from the movement of the protons going through it to attach a phosphate group to an ADP molecule, converting it to ATP. The ATP is then used to supply the rest of the cell with it's energy needs.


This is the end of the line for the glucose molecule. All of the energy stored in it's bonds has been transferred (directly or indirectly) into the chemical bonds of approximately thirty ATP molecules. That's a net gain, since some ATP molecules were consumed in these processes. Most of those thirty ATP molecules were created in stage four, using the electron transport chain.
The original "high energy" C-C and C-H bonds in glucose have been rearranged into the "low energy" C-O and H-O bonds in water and carbon dioxide. These are considered waste products as far as respiration is concerned, because the chemical bonds in water and carbon dioxide are at their lowest (most stable) energy levels due to the high electronegativity of oxygen. No more energy can be squeezed out because transferring hydrogen or carbon from oxygen to any other biologically active atom is an uphill (energy consuming) process. They can, however, be made into energy-rich carbohydrates again through photosynthesis, using the power of the sun to re-create the C-H and C-C bonds and start the photosynthesis/respiration cycle all over again.
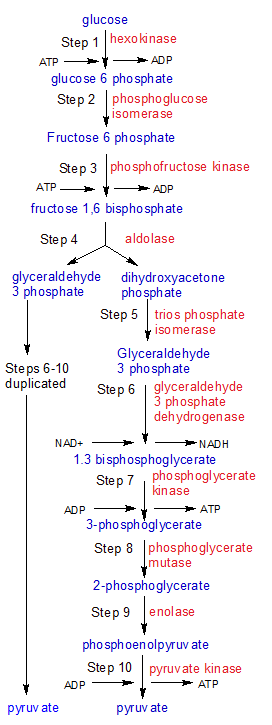
The flowchart above maps out the ten steps of glycolysis. One molecule of glucose enters glycolysis, and two molecules of pyruvate leave it. Note that steps 6-10 are completed twice because two molecules of G3P were created in steps 4 and 5. The substrates are blue and the enzymes are red.
|
STEP 1: Acetyl-CoA (not shown) donates it's acetyl group to an oxaloacetate molecule in a condensation reaction that creates a molecule of citrate. One molecule of water is consumed. The CoA molecule floats away to fetch another acetyl group. We are left with a 6-carbon molecule of citrate, which takes us into step two.
STEP 2: Aconitase converts citrate into isocitrate using a dehydration reaction that moves the hydroxyl group (OH) from the 3rd carbon to the 4th carbon.
STEP 3: Isocitrate dehydrogenase converts isocitrate into alpha ketoglutarate by severing a C-C bond in isocitrate. This discards a CO2 molecule in a reaction called oxidative decarboxylation. The discarded carbon atom trades it's freed-up C-C bond for a C=O double bond in the new CO2, which releases free energy. This energy is used to produce a molecule of NADH from NAD+ through a hydride (H-) transfer to NAD+.
STEP 4: Oxidative decarboxylation is used again to discard another molecule of CO2 and create another molecule of NADH. The 5-carbon alpha ketoglutarate is converted into a 4-carbon succinyl group, which then uses some of the energy released by the casting off of CO2 to form a C-S bond with a CoA molecule, producing succinyl-CoA.
STEP 5: The energy stored in the C-S bond in step 4 is now utilized by the succinyl-CoA synthetase enzyme in step 5. The C-S bond is broken and the CoA is discarded. This leaves us with succinate and some released energy. The energy is used to attach a phosphate group to a molecule of GDP, which produces GTP.
STEP 6: Succinate dehydrogenase oxidizes succinate, converting two C-H bonds and a C-C bond into a C=C double bond. This changes succinate into fumarate and some free energy. The energy is used to attach the two discarded H atoms to a molecule of FAD, producing the energy carrier FADH2.
STEP 7: The fumerase enzyme converts fumerate into malate in a hydrolysis reaction that converts the C=C double bond created in the last step back into a C-C single bond. It does this by splitting a water molecule into OH- and H+ groups and attaching each of them to one of the two middle carbon atoms on fumerate. This forces the two carbons to reduce their double bond to a single bond. Since a C-C bond has been replaced with a C-O bond, there is a release of free energy.
STEP 8: In the final step of the Citric Acid Cycle, the malate dehydrogenase enzyme oxidizes malate into oxaloacetate. The C-O single bond on one of malate's four carbons is converted into a C=O double bond by severing the related C-H and O-H bonds and transferring the two hydrogen atoms to a molecule of NAD+. This creates another energy-carrier molecule of NADH. NADH only needs one hydrogen atom, but both of the electrons, so the extra hydrogen nucleus (H+) is discarded.
Most of the energy in glucose is transferred to ATP in the 4th stage of cellular respiration that's called "oxidative phosphorylation". The mitochondria use this energy (now in the form of NADH) to force protons across it's inner membrane. The crowded protons then diffuse back across the membrane through an opening in ATP synthase, producing ATP in the process. This is how the energy in our food is converted into a form that our cells can use to maintain itself. But this isn't the only possible fate for the energy stored in glucose. It can be "wasted" in thermogenesis.

Thermogenesis bypasses the production of ATP by setting up a detour for the protons as they move back into the mitochondria's interior. Instead of using ATP Synthase, the protons diffuse across the membrane through a protein called thermogenin. The energy is released as heat instead of being used to create ATP.
If converting energy from glucose into heat sounds like a waste of energy, that's because it often is. But in some situations, like when we are infants, it can be useful. We are born without the ability to put on a coat or move towards a source of heat. We haven't even learned how to shiver yet, which is how most people generate heat when they are cold.
Fortunately for the infant, it's born with a large number of specialized fat cells called "brown adipose tissue". These fat cells are different than the normal "white adipose" fat cells because they contain a larger number of mitochondria than the white fat cells. These aren't normal mitochondria either, because they contain lots of the thermogenin proteins. These brown fat cells are designed to burn energy, not store it. This comes in handy for the infant, since they help it stay warm.
When we get older and learn to keep warm in other ways, the brown fat cells decrease in number. Some people lose more brown cells than others, which can influence how much fat their body will store when they are adults. Some people can eat whatever they want and stay thin, while other people eat less and still put on weight. While exercise and other factors like genetics and the metabolic rate contribute to a person's weight, the brown adipose cells also have an impact.