Garden and Plate
The Molecular Biology of Nutrition




The amount of fructose in our diet has increased significantly since we discovered how to separate sucrose (table sugar) from plants like sugar beets and sugar cane. Sucrose is fifty percent fructose and fifty percent glucose. |
Although many of the same molecules and reactions are used when metabolizing glucose and fructose, there are some significant differences in how these two sugars are processed. The most important difference is that fructose can't participate in glycolysis, while glucose can. Most of the cells in our bodies use an enzyme called hexokinase to start the process of glycolysis by phosphorylating glucose. But hexokinase only works on sugars (like glucose) that have a 6-sided ring structure. Fructose has a 5-sided ring structure, so hexokinase can't phosphorylate it. This means that most of the cells in our body can't process fructose. For this reason fructose is metabolized almost exclusively in the liver in a process called fructolysis.
Liver cells (hepatocytes) can process fructose because they contain an enzyme that most other cells don't have. This enzyme is called fructokinase. Fructolysis starts with the phosphorylation of fructose by fructokinase. A few steps later fructolysis produces the same intermediate molecules (GAP and DHAP) that are produced from glucose via glycolysis. But since GAP and DHAP both have phosphate groups attached, they can't leave the liver cell to be used elsewhere. The liver cell can burn some of them for energy, the same as other cells, but it can only use so much.
Since the rest of the body can't process fructose, the liver cells get overwhelmed in a high-fructose diet. The first thing the liver cell does with the excess fructose is store it as glycogen. This is an optimal solution, since glycogen can be broken down into glucose and sent to other cells in the body for their use. But a liver cell can only store a limited amount of glycogen. If a diet is high in fructose the liver runs out of space. When it has all of the glycogen it can store, it uses the extra fructose to create triglycerides, which can be moved outside of the liver for long-term storage as fat. If a person's diet is consistently high in fructose, the excess fat never gets used, and builds up over time. While excess glucose can also be stored as fat, there are unique health concerns with fructose-produced fatty-acids. Fat produced in the liver isn't regulated or stored in the same way as the fat from glucose.
|
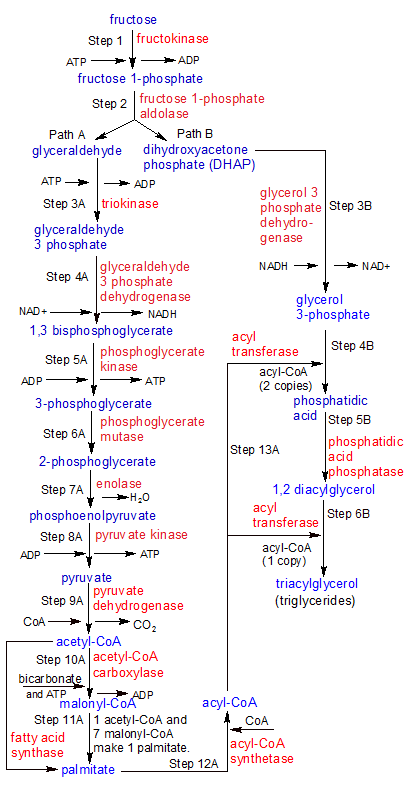
Step 1: Fructolysis starts when liver cells (hepatocytes) use the fructokinase enzyme to convert fructose into fructose 1-phosphate. Fructokinase transfers a phosphate group from ATP to fructose, producing fructose 1-phosphate.
Step 2: Fructose 1-phosphate aldolase splits fructose 1-phosphate into two molecules: glyceraldehyde and dihydroxyacetone (DHAP). These two molecules take separate reaction pathways, so the flowchart splits after step two. We'll look at Path A first, then at Path B. Note that the products of Path A eventually feed back into Path B.
Step 3A: Triokinase transfers a phosphate group from ATP to glyceraldehyde, producing a molecule of glyceraldehyde 3-phosphate (G3P).
Metabolic Pathway Notes: At this point the liver cell can break G3P down for it's energy, using the same glycolysis steps (6-10) that it uses for glucose. What it can't do is transfer any excess G3P to any other cells, because phosphorylated molecules like G3P can't cross the cell membrane. Since the liver has to process all of the fructose itself, it gets far more than it can use for it's own energy needs. So instead of burning it for energy, it stores it as glycogen in the liver. When it has plenty of glycogen, it will then use it to create triglycerides. In this flowchart we'll follow the steps used to create triglycerides. See the glycolysis flowchart to see how glyceraldehyde 3 phosphate is used to create ATP for immediate energy use.
Step 4A: Glyceraldehyde 3-phosphate dehydrogenase uses energy stored in NAD+ to add a 2nd phosphate group to glyceraldehyde-3 phosphate, creating 1,3 bisphosphoglycerate. NAD+ was converted into NADH in the process. An unattached molecule of inorganic phosphate was used, so no ATP was needed.
Step 5A: Phosphoglycerate kinase removes a phosphate group from carbon-1 on 1,3 bisphosphoglycerate, converting it into 3-phosphoglycerate. A molecule of ATP is created in the process.
Step 6A: Phosphoglycerate mutase moves the phosphate group from carbon-3 to carbon-2, converting 3-phosphoglycerate into 2-phosphoglycerate.
Step 7A: Enolase converts 2-phosphoglycerate into phosphoenolpyruvate in a dehydration reaction that discards a molecule of water.
Step 8A: Pyruvate kinase de-phosphorylates phosphoenolpyruvate, creating ATP and pyruvate.
Step 9A: Pyruvate dehydrogenase removes a CO2 molecule from pyruvate, converting it into a two-carbon acetyl group. The same enzyme then attaches the acetyl group to CoA, creating acetyl-CoA.
Step 10A: Acetyl-CoA carboxylase uses energy from ATP to combine bicarbonate with acetyl-CoA to create malonyl-CoA. Both acetyl-CoA and malonyl-CoA have two carbon acetyl groups attached to them. These groups will provide the substance of the fatty acid chains.
Step 11A: Fatty acid synthase uses a 4-step process to transfer the two-carbon acetyl groups from seven malonyl-CoA molecules, attaching them (one after another) to the two-carbon acetyl group on the end of a single acetyl-CoA molecule. This process is repeated until the acetyl group on acetyl-CoA is 16 carbons long. At this point, the 16-carbon long string separates from the newly formed acyl-CoA to become palmitate, a free fatty acid. A molecule of CoA also remains, to be used again.
Step 12A: Acyl-CoA synthetase binds palmitate to CoA, creating acyl-CoA. This is identical to the acyl-CoA molecule we had near the end of step 11A, before palmitate separated from CoA. Instead of separating in step 11A, they could have skipped this step and gone on to step 13A as acyl-CoA. Step 12A is only needed if you have unattached palmitate and CoA floating around.
Step 13A: This step feeds into steps 4B and 6B of Path B in our metabolic pathway. Acyl transferase transfers the 16-carbon palmitate strands from two acyl-CoA molecules to one glycerol 3-phosphate molecule, giving it two of the three strands it needs in order to be a triglyceride. The same enzyme transfers a third strand of palmitate to the same molecule in step 6B, after it's been converted to 1,2 diacylglyceral. For more details on how all of these parts come together to create a triglyceride, see the Path B steps below.
|
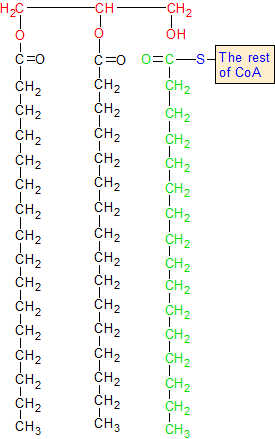
Note: All of these CoA molecules are confusing, so let me clear this up. We started Step 11A with a molecule of malonyl-CoA, and ended Step 12A with a molecule of acyl-CoA. It should be noted here that CoA (coenzyme A) is used in these steps as a carrier for two-carbon groups that take various forms. Acetyl-CoA is CoA with a single two-carbon acetyl group attached. Acyl-CoA is acetyl-CoA with eight two-carbon groups attached (The one it had originally as acetyl-CoA, and seven donated by seven different malonyl-CoA molecules). Malonyl-CoA is acetyl-CoA with bicarbonate added to it. Acetyl-CoA becomes acyl-CoA when it aquires the seven additional acetyl groups. This string of eight two-carbon acetyl groups becomes palmitate when it separates from CoA.
Step 3B: Glycerol 3 phosphate dehydrogenase uses H- from an NADH molecule and a free floating proton H+ to convert DHAP into glycerol 3-phosphate. It does this by transforming the C=O double bond on the center carbon of the 3-carbon molecule to a C-O single bond. One Hydrogen atom binds to the carbon, and the other to the oxygen atom. Now the oxygen atoms on both carbon-1 and carbon-2 in the glycerol 3-phosphate molecule have an O-H single bond that can be used to attach a palmitate strand.
Step 4B: Acyl transferase transfers palmitate from acyl-CoA to each of the two oxygens on glycerol 3-phosphate that are connected to carbons 1 and 2 (Carbon-3 is inaccessible because it's attached to a phosphate group). The O- ion on the end of each palmitate combines with each of the two OH groups on glycerol 3-phosphate, discarding an OH- hydroxyl group in the process. A molecule of phosphatidic acid is produced.
Step 5B: Two of glycerol 3-phosphate's three carbon atoms are now attached to palmitate, but the 3rd carbon is attached to a phosphate group. Phosphatidic acid phosphatase de-phosphoylates glycerol 3-phosphate, converting it into 1,2 diacylglycerol.
Step 6B: With the 3rd carbon in 1,2 diacylglycerol now attached to an OH group instead of a phosphate group, the same acyl transferase enzyme that transferred palmitate strands from acyl-CoA to carbons 1 and 2 in step 4B can now transfer a third strand to carbon-3. This completes the creation of the triacylglycerol molecule (a triglyceride). The conversion of fructose into a triglyceride (carbohydrate into fat) is now complete. And because the triglyceride has no phosphate groups attached, it can be transferred out of the liver cell for long-term storage elsewhere.